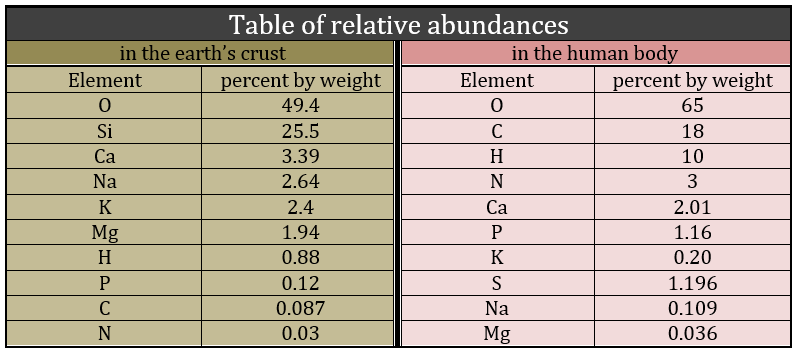
La biochimie est un domaine de la chimie lié aux corps vivants, animaux ou végétaux. Ce champ est loin de la chimie inorganique et une simple comparaison peut le montrer: la répartition des molécules dans la croûte terrestre et dans un corps vivant est totalement différente. Le premier est essentiellement composé de silice et le second de carbone.
Historiquement, la première théorie sur la vie était qu’elle est donnée par Dieu et seulement par Lui. Les molécules sont inanimées et ne peuvent pas être transformées en un corps vivant sauf si Dieu y insuffle la vie. C’est ce qu’on appelle la théorie du vitalisme.
Miller a montré à travers ses expériences qu’il était possible de créer des molécules appartenant à des corps vivants avec des molécules simples. Les expériences de Miller consistent dans la modélisation des conditions sur Terre avant l’apparition de la vie. Il n’y avait pas beaucoup d’oxygène en ce moment et l’atmosphère était essentiellement composée d’ammoniac, de méthane, d’hydrogène et d’eau. La température était bien plus élevée et il y avait beaucoup d’éclairs. Dans ces conditions, certaines molécules présentes dans les organismes vivants se sont formées spontanément: urée, glycine, alanine, …
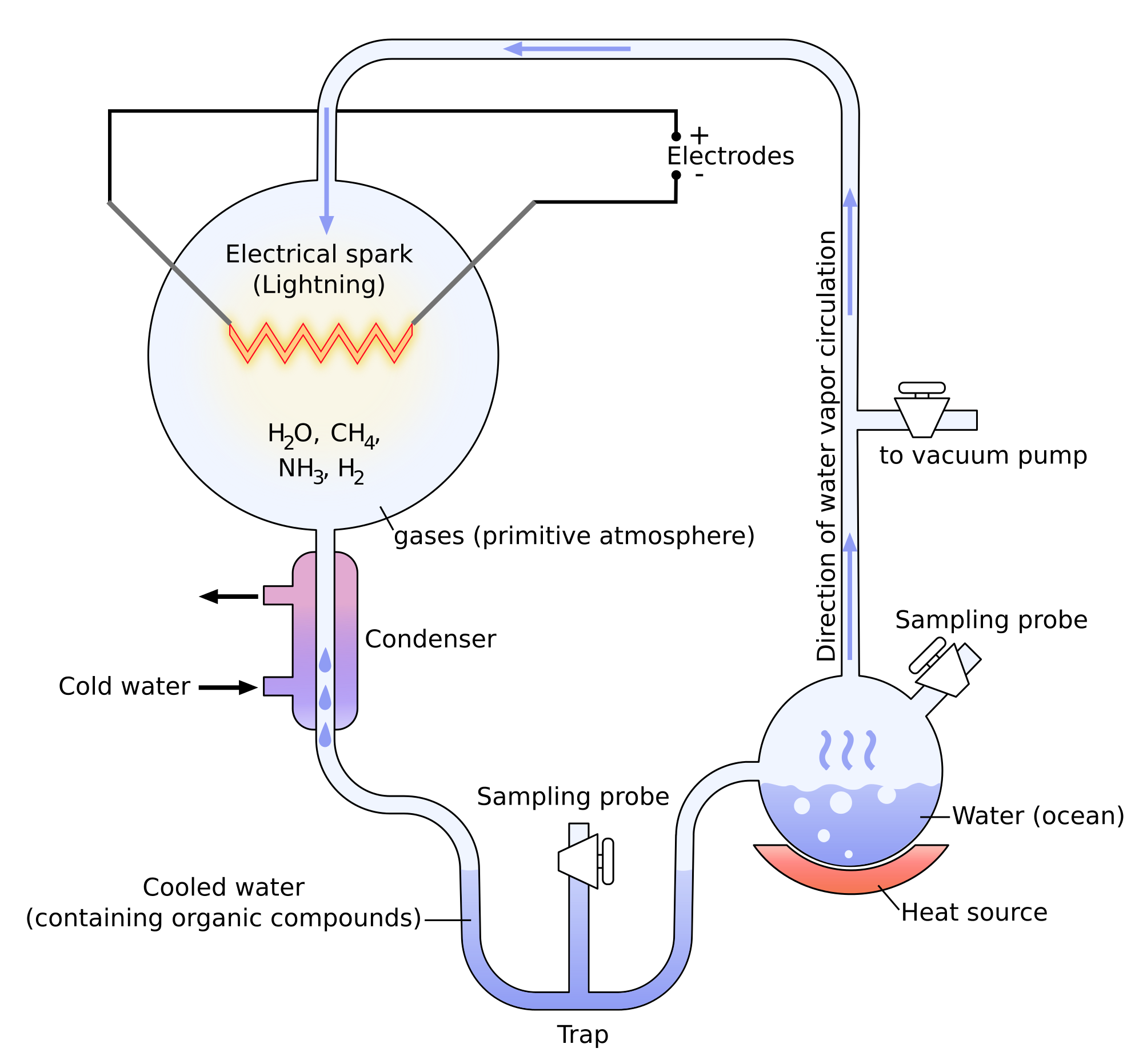
Pourtant, il n’y avait aucune trace de protéines et surtout d’ARN (acides ribonucléiques), les molécules capables d’écrire des molécules d’ADN (acides désoxyribonucléiques). Il a été montré plus tard que certaines molécules de météorites permettaient la formation d’ARN.
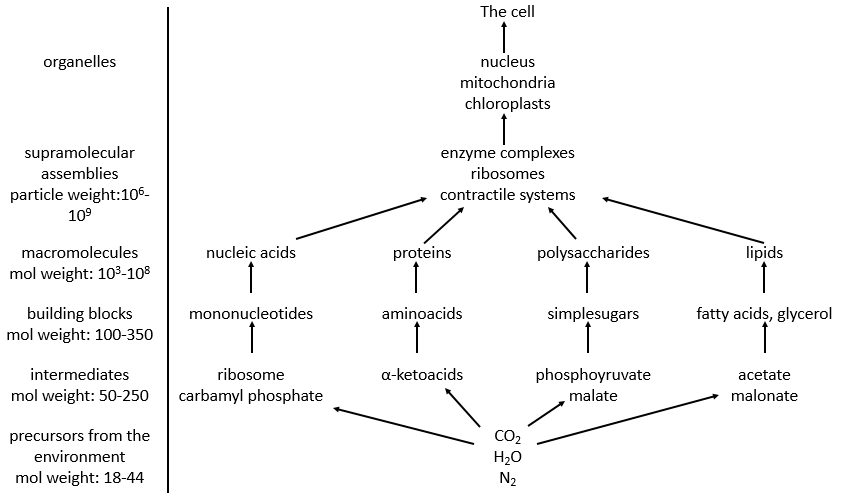
Principales caractéristiques de la biochimie
Les limitations
Parmi les bonnes centaines d’atomes du tableau de Mendeleev, on ne trouve que 16 liaisons différentes en biochimie mais elle permet la formation d’une infinité de molécules organiques. Ceci est d’autant plus surprenant que les réactions impliquées dans les processus biologiques sont limitées par les conditions biologiques de l’existence: il n’y a aucune manière qu’une réaction qui nécessite une température de 100 ° C ait lieu dans notre corps. Cela endommagerait les molécules environnantes et brûlerait les tissus. Comme toute réaction chimique, ils doivent obéir aux règles de la thermodynamique et trouver l’énergie nécessaire pour faire la réaction ailleurs.
La spécialisation
Dans un corps, les réactions sont faites dans des cellules et peuvent également être compartimentées à l’intérieur de la cellule. Toutes les cellules ne peuvent pas faire toutes les réactions et certaines cellules sont spécialisées. La spécialisation est écrite dans l’information génétique et se traduit par la présence ou l’absence de certaines enzymes. De plus, dans une cellule, il n’y a pas le même type de réactions dans les mitochondries que dans le réticulum endoplasmique. C’est parce que les réactions sont régulées, positivement ou négativement, par des catalyseurs.
Les Enzymes
La température doit rester basse (~ 37 ° C) et ne peut pas beaucoup varier. ΔG = ΔH-TΔS doit être négatif et proche de zéro. La vitesse de réaction serait proche de zéro sans catalyseur.
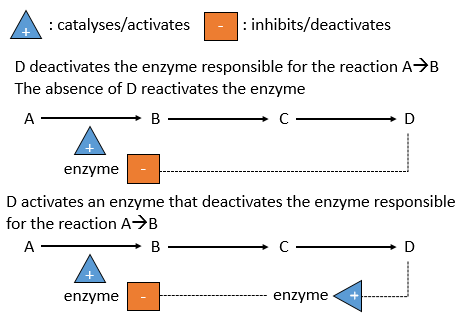
Il y a un système de régulation dans les cellules: l’activité des catalyseurs dépend des besoins de la cellule. Si la cellule est pleine d’une espèce D, il n’est pas nécessaire d’en accumuler davantage alors que les ressources pourraient être utilisées dans un processus différent. La présence de l’espèce D influencera un catalyseur (souvent une enzyme) qui agit sur une réaction conduisant à la formation de D. La présence de D peut inhiber une enzyme responsable de la réaction ou activer une enzyme inhibant la réaction. Au contraire, l’absence de D peut également être détectée par cette enzyme ou par une autre pour favoriser la formation de D.
La gestion de l’énergie est un problème global pour le corps: les processus ne peuvent pas consommer ou rejeter trop de chaleur à la fois. Il pourrait endommager les cellules environnantes et inhiber la plupart des enzymes: la température du corps est habituellement dans la gamme optimale d’efficacité pour les enzymes et s’il y a une variation de température, l’activité des enzymes diminue de manière significative.
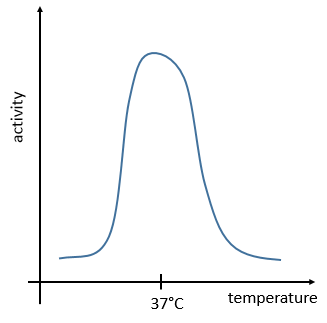
La régulation
De plus, il y a des périodes durant lesquelles nous consommons moins d’énergie (quand nous dormons par exemple) et des périodes où le corps a besoin de plus de ressources (en faisant du sport, en période de stress, …). Ces périodes ne coïncident pas avec les périodes pendant lesquelles nous produisons l’énergie (quand nous mangeons). Il doit donc y avoir un moyen efficace de stocker l’énergie quand elle est produite et de la libérer en cas de besoin. Cela se fait par la formation d’ATP (adénosine triphosphate) à partir d’ADP (adénosine diphosphate). Cette réaction n’est possible que dans les mitochondries mais l’énergie est nécessaire partout. L’énergie doit donc également être transportée dans les différents compartiments des cellules et dans le corps.
Ce cours se concentrera sur ces problèmes à travers l’exemple de la dégradation du glucose. Avant cela, nous allons faire une introduction des principaux types de molécules que nous trouvons le corps.
Les molécules du corps :
Ce cours se concentrera sur ces problèmes à travers l’exemple de la dégradation du glucose. Avant cela, nous allons faire une introduction des principaux types de molécules que nous trouvons le corps.
L’ eau
Notre corps est composé à 60-70% d’eau. La plupart des réactions sont ainsi réalisées dans des conditions aqueuses et polaires. L’eau implique une organisation entre les molécules. Dans la glace, une molécule d’eau forme des liaisons 4H avec d’autres molécules H2O.
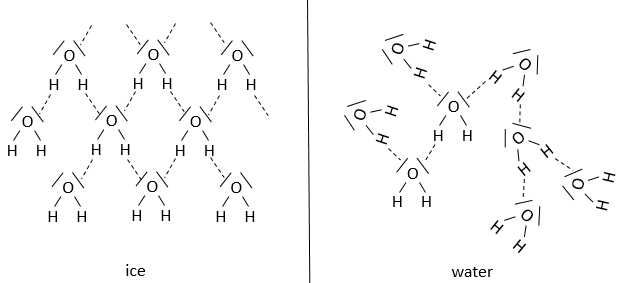
À la température du corps, ce nombre chute à environ 3,4. Les liaisons H se rompent facilement et fréquemment: leur énergie est de ~ 4,5 kcal / mol (contre ~ 110 kcal / mol pour une liaison covalente) et leur durée de vie est d’environ 10-9s.
Les liaisons H lient les molécules d’eau ensemble mais aussi la molécule d’eau à tout atome électronégatif (par exemple R-OH, R-CO-R, R-COOH, R-NH2 …). Les groupes qui forment des liaisons H sont appelés groupes hydrophiles. Les groupes qui ne permettent pas la formation de telles liaisons sont appelés groupes hydrophobes (une chaîne aliphatique par exemple, un phényle …). Les premières sont solubles dans l’eau alors que les secondes ne le sont pas (elles forment deux phases séparées). Une molécule qui porte des groupes hydrophiles et hydrophobes est appelée amphipathique.
La transpiration est un moyen que le corps a trouvé pour réguler la température: l’énergie de vaporisation de l’eau est de 540cal / g d’eau. Pour compenser la hausse de température de 1 degré, 2 grammes d’eau sont évaporés.
Les lipides :
Les lipides sont un groupe de molécules naturelles composées habituellement d’une tête hydrophile et d’une ou plusieurs chaînes hydrophobes.
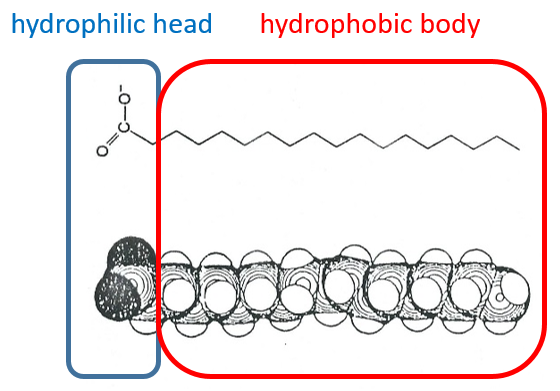
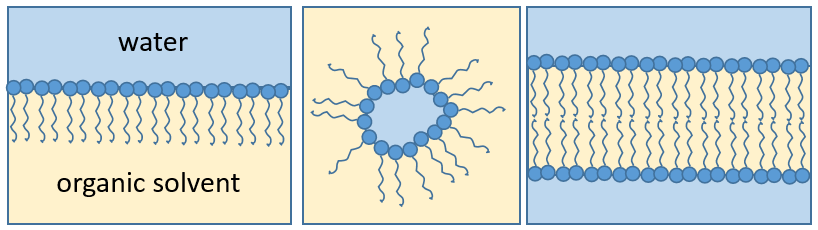
La présence de la tête polaire ne signifie pas que la molécule entière est soluble dans l’eau. Les chaînes hydrophobes sont insolubles dans l’eau et s’agrègent ensemble pour minimiser la surface de contact avec les molécules d’eau. Les principales fonctions biologiques des lipides comprennent le stockage d’énergie, la signalisation et l’action en tant que composants structuraux des membranes cellulaires. Les membranes qu’elles forment peuvent être soit une mono-couche (séparant une phase aqueuse et une phase organique) soit une bi-couche (séparant généralement deux phases aqueuses).
Les lipides sont triés en fonction de la capacité à former du savon. Pour ce faire, le lipide doit contenir un acide gras (une chaîne hydrophobe finissant par un groupe COOH hydrophile).
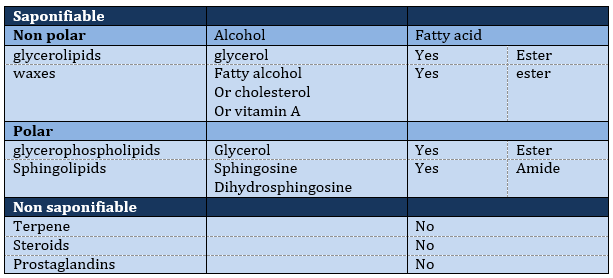
Leur nomenclature est terminée par ~ (an) oate. Par exemple, CH3(CH2)10COO– est appelé dodécanoate. Certains lipides portent des noms provenant du végétal dont ils proviennent. Par exemple CH3(CH2)14COO– est appelé palmitate car il provient de palmiers. S’il y a une double liaison dans la chaîne, sa position est indiquée au début du nom par un Δx où x est la position du premier carbone avec un C = C et le suffixe est maintenant ~ enoate. Par exemple, CH3(CH2)CH=CH(CH2)7COO–CH3 est appelé A9-hexadécénoate.
La présence d’une double liaison durcit la structure du lipide. Un lipide sans C = C est dit saturé et toutes les liaisons peuvent tourner. Ces lipides sont très flexibles. D’autre part, les C = C fixent la conformation localement car la double liaison ne peut pas tourner. En conséquence, la cohésion entre les chaînes est plus complexe et le nombre d’interactions de van der Waals diminue (en d’autres termes, les chaînes ne peuvent pas s’imbiber aussi bien que les chaînes saturées). Un lipide à double liaison est appelé insaturé. S’il y a plusieurs C = C, on parle de polyinsaturés. Pour illustrer la différence que fait un C = C, le tableau suivant montre une série de lipides avec leur température de fusion.
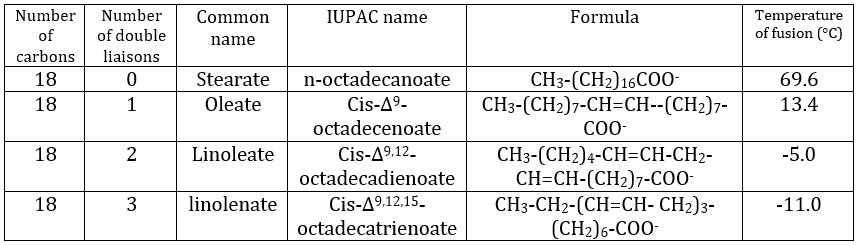
Les lipides insaturés se trouvent sous la forme d’une huile tandis que les lipides saturés se présentent sous la forme de graisses. Il est possible d’hydrogéner les lipides insaturés pour éliminer le C = C.
Lipides saponifiables
Le glycérol est un polyalcool avec 3 groupes -OH sur une chaîne à 3 carbones. Quand il réagit avec 3 équivalents de RCOO- (un lipide), il forme un triacylglycérol. Le carbone noté * C est chiral si R1 et R2 sont différents. La réaction dans la direction opposée est la réaction de saponification:
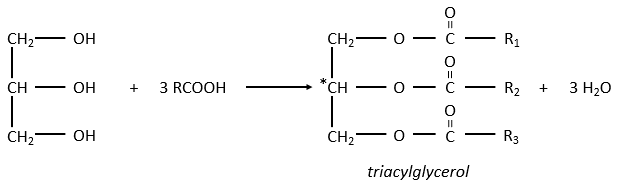
La réaction dans la direction opposée est la réaction de saponification:
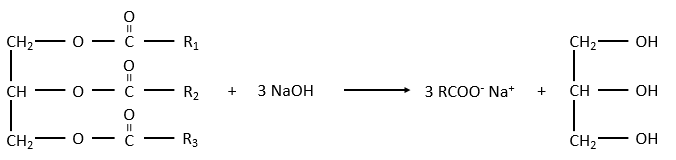
RCOONa est un savon. Le principe est assez simple: le savon forme des micelles autour de la saleté qui n’est pas soluble dans l’eau. Les chaînes hydrophobes encerclent la saleté et les têtes hydrophiles sont en contact avec l’eau.
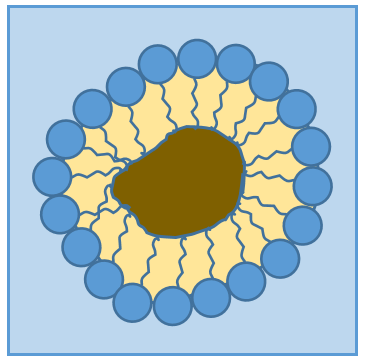
The whole thing is soluble in the water. Lipids are an energetic reservoir stocked under the form of pellets of fat in cells called adipocytes.
Waxes are a combination of a fatty acid with a fatty alcohol.
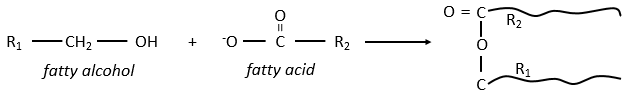
Ils sont très hydrophobes et constituent une protection efficace contre l’humidité (exemple: la laine de mouton) ou pour empêcher l’eau de s’évaporer (ex: plantes tropicales, cactus). Les baleines à dents utilisent un mélange de cires et de triacylglycérol, appelé spermaceti pour nager sans effort dans les profondeurs. A 37 ° C, le mélange est une huile mais la température baisse et le mélange cristallise. Sa densité augmente, ce qui permet au cachalot de flotter à cette profondeur.
Les lipides polaires présentent souvent un phosphate ou une amine chargée
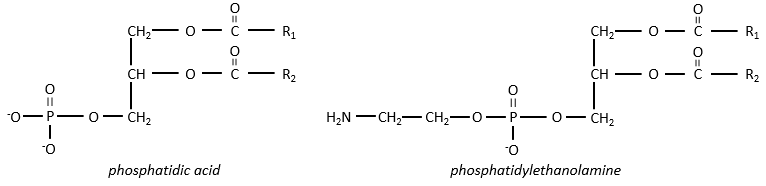
Il y a beaucoup de dérivés de l’acide phosphatidique. Notons l’acide phosphatidique R,
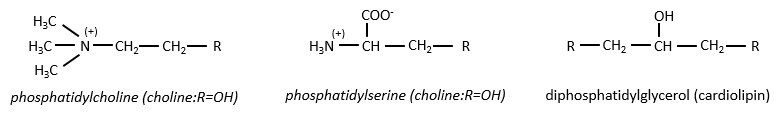
Beaucoup d’entre eux constituent les membranes de la cellule. Les sucres peuvent également être liées aux lipides par liaison ester et servir, dans ce cas, d’élément de signalisation.
Les sphingolipides sont des dérivés de la sphingosine et de la dihydrosphingosine.
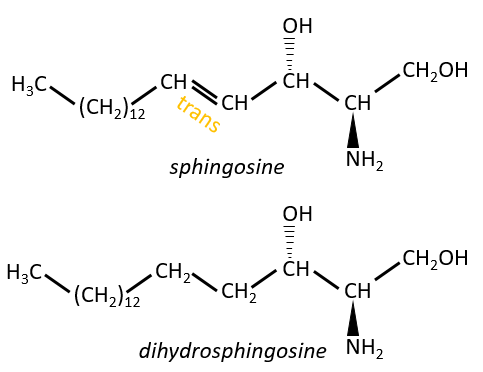
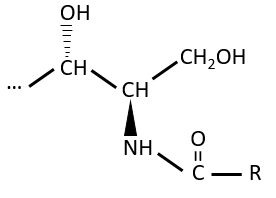
Les lipides sont liés par une liaison amide.
Les éléments donnant des propriétés différentes à la molécule peuvent se lier à l’alcool. Par exemple, une choline peut se lier à l’aide d’une liaison phosphodiester.
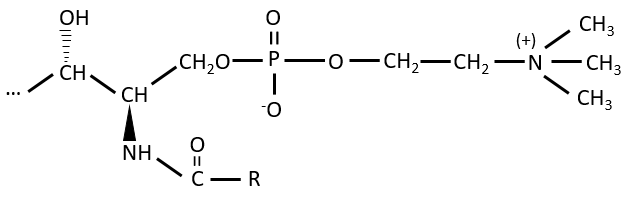
Cette molécule est une sphingomyéline et se trouve dans la membrane qui entoure certains axones des cellules nerveuses. Sur l’ester, un ucre peut être lié. Ces molécules sont appelées glycosphingolipides neutres ou cérébrosides parce qu’elles se trouvent souvent dans les membranes du cerveau. Encore une fois, le sucre est un élément de reconnaissance / signalisation.
Le sucre peut aussi être acide (par exemple un acide sialique) et alors la molécule est un glycosphingolipide acide, servant habituellement de récepteur.
Lipides non saponifiables
Terpènes
Ce sont de petites molécules dérivant de l’isoprène.
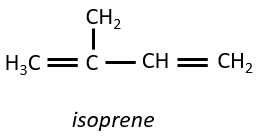
Les plus petits terpènes sont les monoterpènes, résultant de la combinaison de deux isoprènes.
8 équivalents de l’isoprène forment le bêta-carothène. Cette molécule de 40 carbones est un antioxydant qui protège des radicaux libres: elle est facilement excitée car les électrons peuvent être délocalisés.
C’est aussi un précurseur de la vitamine A. C’est un groupe de composés organiques nutritionnels insaturés (trouvés dans les carottes, entre autres) qui comprend le rétinol, le rétinal, l’acide rétinoïque et plusieurs caroténoïdes de provitamine A (notamment le bêta-carotène).
La vitamine A a de multiples fonctions: elle est importante pour la croissance et le développement, pour le maintien du système immunitaire et pour une bonne vision. La rétine de l’œil a besoin de vitamine A sous forme de rétinal, qui se combine avec la protéine opsine pour former la rhodopsine, la molécule absorbant la lumière nécessaire à la fois à la vision faible (vision scotopique) et à la vision des couleurs.
Il y a deux types de capteurs dans l’œil: les cônes et les bâtonnets. Le premier détecte les couleurs et les secondes la lumière. Les bâtonnets sont composées d’une série de disques bilipidiques avec des récepteurs de rhodopsine. Entre les hélices d’opsin est le 11-cis-rétinal
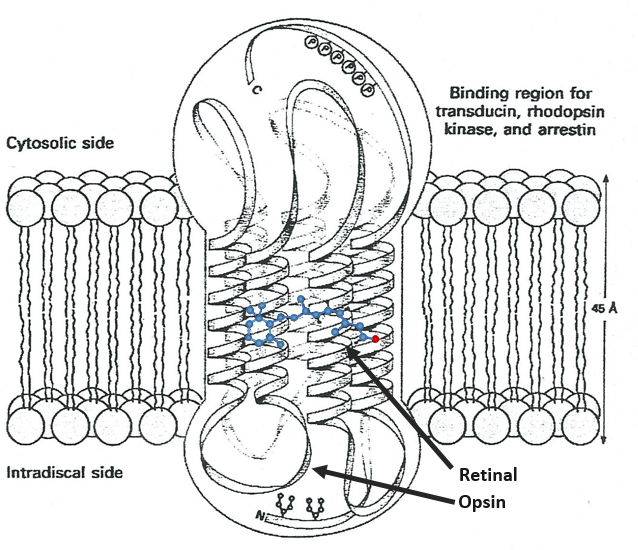
Toutes les liaisons sauf une dans la rétinal sont trans. Quand un photon frappe le rétinal, cette liaison devient trans, perturbant la molécule, puis les hélices protéiques qui transfèrent le signal nerveux. Un photon est suffisant pour générer un signal.
Dans les cônes, il y a des modifications spécifiques dans la structure de la protéine et le photon doit être dans une plage spécifique de longueur d’onde (c’est-à-dire une couleur) pour transférer le signal.
Les stéroïdes :
La base des stéroïdes est le stérol, une molécule faite de trois cycles de 6 C et un de 5 C.
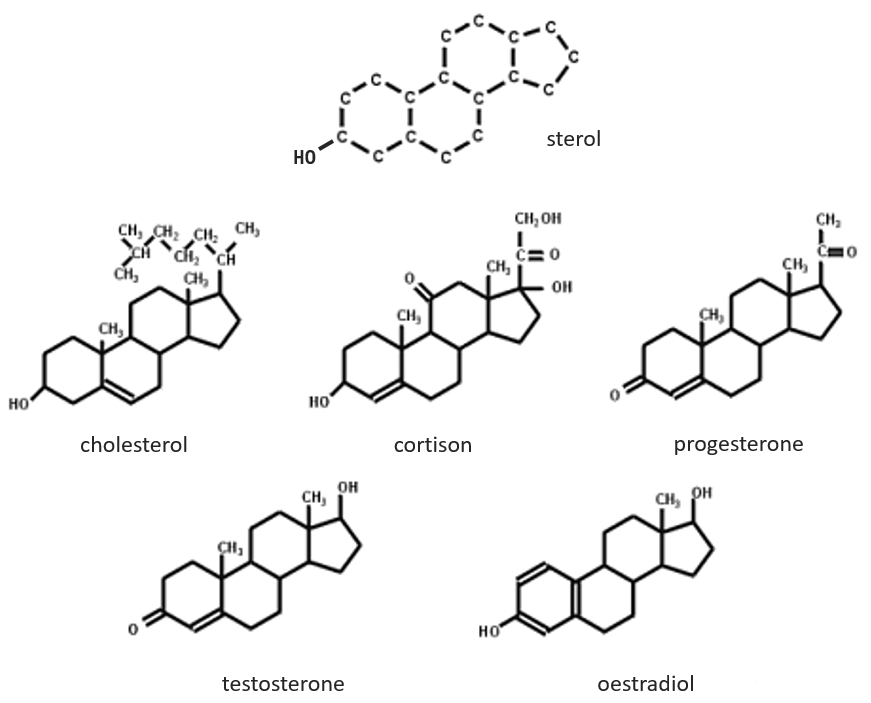
Le cholestérol est complètement hydrophobe. C’est un précurseur de plusieurs hormones, de la vitamine D et du sel biliaire.
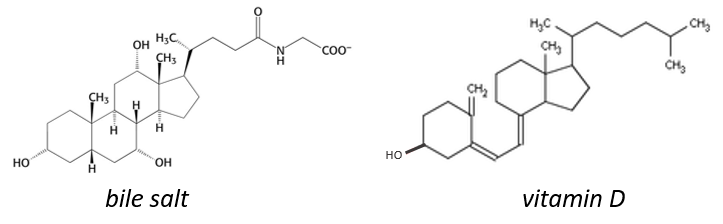
On entend souvent parler du bon et du mauvais cholestérol. C’est en fait une locution incorrecte. Le mode de transport du cholestérol est en fait le principe important. Le cholestérol est transporté par les lipoprotéines (une combinaison de lipides et de protéines) appelées LDL et HDL (pour les lipoprotéines de basse et haute densité). Le LDL transporte le cholestérol du foie vers le corps. Il est lié à certains récepteurs spécifiques sur LDL. Le rôle du HDL est de ramener le cholestérol inutilisé au foie où il est détruit. Il y a donc un équilibre entre le cholestérol qui quitte le foie et qui revient au foie.
Les prostaglandines :
Ce sont des hormones agissant sur une courte distance, avec une durée de vie courte et elles sont toutes fabriquées à partir de l’arachidonate, un lipide de la membrane cellulaire, par une phospholipase. Les prostaglandines activent de nombreux récepteurs membranaires.
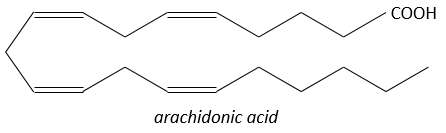
A la suite d’un stimulus donné, certains agents sont activés pour activer la PLA2 (phospholipase A2)PLA2. La PLA2 clive les phospholipides de la membrane pour obtenir des acides gras. A partir d’un acide gras 4 types de prostaglandines sont formés:
1) prostacycline: ils se trouvent dans la paroi des vaisseaux sanguins et empêchent la coagulation.
2) PGE et PGF (Prostaglandines E et F): elles sont responsables du cycle du sommeil et des contractions à l’accouchement.
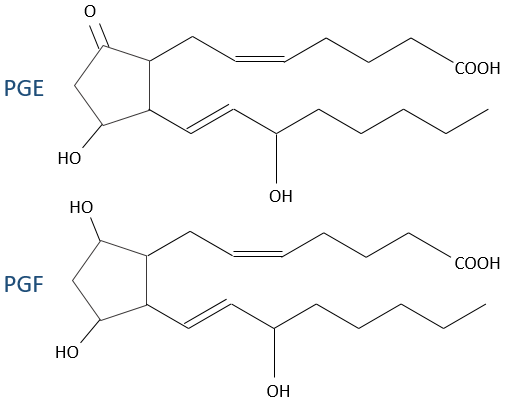
- thromboxane: ils sont impliqués dans le processus de coagulation.

- leucotriènes: nous les trouvons dans la paroi des poumons. Les asthmatiques ont une surproduction de leucotriènes.
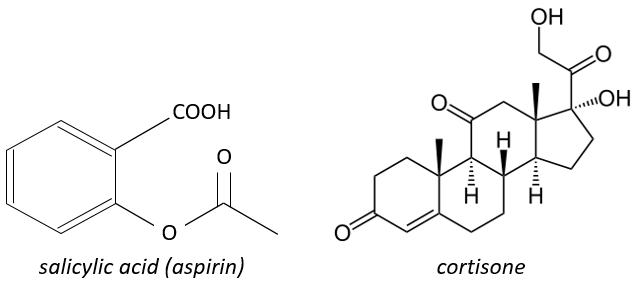
L’aspirine bloque l’activité/production de la cyclo-oxygénase qui est une enzyme responsable de la formation des quatre types de prostaglandines à partir de l’arachidonate. La cortisone bloque la PLA2.
Fonction principale des lipides: constituant des membranes
Les membranes des cellules séparent l’intérieur de l’extérieur des cellules. Les deux côtés sont aqueux mais l’intérieur de la membrane est hydrophobe. Les membranes bilipidiques se forment spontanément (appelées liposomes). Les membranes sont composées de lipides et de protéines dont la composition dépend de la cellule: essentiellement, les protéines permettent les échanges et les transports d’un côté de la membrane à l’autre tandis que les lipides sont la paroi en béton de la membrane. Le cholestérol se place dans la partie hydrophobe de la membrane pour la consolider.